
The Eternal World War Of The Underground
The brutal world of ant warfare, the global battlefield and which species are winning

When I looked at writing an article about ant warfare I started gathering research notes, before realising I could fill several books. Therefore to keep this a humane length I have opted to write an article roughly in the style of a military intelligence brief, hopefully this has kept the details swift and to the point, without losing breadth and depth. If its not to your liking, consider it a lone experiment.
Along one boundary in the coastal region designated “San Diego, California,” approximately thirty million combatants die every year. The front line has not moved appreciably in a decade. In the grimdark of the near future, there is only war and more war.
EXECUTIVE SUMMARY
An armed conflict of planetary scope has been ongoing for approximately forty million years. It currently spans six continents, involves at minimum six distinct invasive military powers and shows no sign of resolution. The primary combatants are eusocial insects of the family Formicidae (vulgar - ants), with approximately 25,000 known species, of which a small number have achieved global force projection through a combination of chemical weapons, slave armies, suicide units and accidental alliances with the planet’s dominant hominid species via its cargo shipping networks.
The latter population is largely unaware of the conflict. Where they are aware, they are still losing.
This briefing covers: (1) the general military capabilities of the combatants (2) the campaign history and current disposition of the conflict’s dominant power (3) the secondary invasions complicating that picture (4) an assessment of likely future trajectory.
PART ONE: CAPABILITIES ASSESSMENT
Survey of belligerent armaments, doctrines, special units
1.1 COMBAT DOCTRINE
Ant military doctrine ranges from ritualised deterrence to industrial-scale annihilation. The variance is significant, we impress upon the reader that these are far from a unified force.
Deterrence model. Myrmecocystus mimicus (the honeypot ant):
Colonies of 20–130 workers deploy hundreds of soldiers for pre-engagement assessment
Combatants perform stereotyped threat displays: stilted locomotion, raised abdomens, antennal percussion
Casualty rate during tournaments: near zero
Purpose: force assessment before committing to decisive action
When imbalance is established, the weaker colony is raided and enslaved
Assessment: Cold War logic. Show of force followed, when asymmetry is confirmed, by annexation.
Human-shield doctrine. Pheidologeton diversus (marauder ant):
Minor workers (expendable, low-cost) deployed for frontline mass
Major workers (high-cost, high-capability) held in reserve until breach is achieved
Colony displays no apparent grief response to minor worker losses
Assessment: Identical in structure to ancient conscript-and-elite armies. Independently derived.
Age-stratified deployment. Solenopsis invicta (fire ant):
Months-old workers: assigned to combat. Weeks-old workers: assigned to flight/withdrawal
Days-old workers: feign death upon contact
Source: Cassill (2008), Naturwissenschaften
Assessment: A triage doctrine, the colony produces its own order of battle.
The mathematics of mass: Champer & Schlenoff (2024) confirmed that ant battles obey Lanchester’s Square Law. This principle was originally derived for WW1 aerial combat, it states that in open-terrain engagements, doubling your forces doesn’t simply double your advantage, but rather quadruples it. Numerical superiority compounds exponentially. Lanchester’s Law is arguably the dominant point of ant success worldwide.



1.2 CHEMICAL WEAPONS PROGRAMME
The Formicidae chemical arsenal is the most sophisticated battlefield pharmacology on the planet. The following represents a partial inventory.
Primary weapon systems:
Formic acid (subfamily Formicinae): Sprayed via specialised acidopore gland. Formica sanguinea (blood-red ant) uses a two-stage system, first Dufour’s gland compounds (hendecane, decylacetate, dodecylacetate) dissolve the enemy cuticle first, then the acid penetrates the exposed tracheal system. Assessment: shell, then gas.
Solenopsin (fire ant, Solenopsis invicta): Piperidine alkaloid. Venom composition >95% alkaloid by mass. Cytotoxic, hemolytic, fungicidal, insecticidal. LD50 data: Pogonomyrmex harvester ants are 684× more resistant than Argentine ants. Fire ants are 330× more resistant than Argentine ants to their own weapon. Assessment: armed and armoured against their own ordnance.
Propaganda substances (Formica subintegra, parasitic slave-making ant): Hypertrophied Dufour’s glands contain ~700μg of decyl acetate, dodecyl acetate, and tetradecyl acetate. During raids, this mixture is discharged at any defending workers which triggers the target colony’s own alarm pheromones. Defenders then interpret their nestmates as threats leading to widespread colony panic. (Regnier & Wilson 1971)
This propaganda weapon is almost certainly unique in the biological world, nothing else in this conflict or arguably in the broader study of warfare, quite matches it. The attacking force reprograms the enemy’s communications network, and the defenders route themselves. Regnier and Wilson called these chemicals “propaganda substances”, they remain one of the most elegant methods of warfare in any species.
Defensive countermeasures:
Tawny crazy ant chemical antidote (Nylanderia fulva, a.k.a Raspberry crazy ant or Caribbean crazy ant): When contaminated with fire ant solenopsin, workers apply self-produced formic acid to their own bodies. Survival rate with treatment: 98%. Without: 48%. (LeBrun et al. 2014). This is the first documented instance of any insect producing and self-applying a chemical detoxification agent in combat conditions. It was derived from an evolutionary arms race conducted over millions of years in South America and was imported, inadvertently, to a theatre where fire ants had held dominance for eighty years.
1.3 SPECIAL UNITS
Trap-jaw ant (Odontomachus bauri):
Mandible closure speed: 35.5–64.3 m/s (up to 230 km/h)
Strike duration: 0.13 milliseconds. Peak acceleration: ~100,000 g
Mechanism: latch-release of stored elastic energy; trigger-hair contact initiates strike
Secondary function: mandible strike against hard surface launches ant into air becomes an escape mechanism
Designated “probably the fastest reflex yet described for any animal” (Gronenberg 1995)
Assessment: a frontline unit with its own ejection system.
Exploding ants (Colobopsis explodens, described Laciny et al., 2018):
Autothysis is the voluntary gaster rupture under muscular contraction
Explosion releases sticky, toxic, antimicrobial secretion from glands extending the length of the body. Kills or immobilises attackers and disinfects the immediate area
Assessment: the suicide vest which also sanitises the crater, just a standard operating procedure for this unit.
Phragmotic doorkeepers (Colobopsis spp.):
Soldier ants possess flattened, shield-shaped heads
Function: physically block nest entrance with cranium, requires significant force to dislodge or displace
Assessment: specialised caste as living door. They are also the fallback position, in the event of a breach, the door can become a fighter.



Megaponera analis, the field hospital:
Megaponera analis (Matabele ants) raids termite colonies in organised columns of up to 600 workers. Termite soldiers fight back, many ants suffer injuries (legs seized, limbs locked, bodies damaged). The injured ant releases dimethyl disulfide (DMDS) and dimethyl trisulfide (DMTS), which is a chemical distress signal. Uninjured nestmates carry the wounded back to the nest.
Workers apply metapleural gland secretions to the injuries. Those secretions, analysed by Frank et al (2023), contain 112 antimicrobial compounds and 41 proteins. Without treatment: 80% of injured ants die. With treatment: 10%. Colony populations are approximately 29% larger as a direct result of this practice.
Megaponera analis is the only known non-human animal, besides chimpanzees, to treat infected wounds with what are functionally antibiotics. The colony derived this practice from evolutionary pressure, preceding human medicine by tens of millions of years.
1.4 SLAVE ARMIES AND OCCUPIED FORCES
Dulosis is the term of the raiding and enslavement of foreign colonies. This practice has evolved independently approximately ten times across roughly sixty known species, distributed across multiple genera. First documented by Pierre Huber in 1810.
Polyergus spp. (Amazon ants), the professional raider:
Mandibles: sickle-shaped, useless for any purpose except combat and larval transport
Colony composition: 3–5 slave-maker workers per 70–90% enslaved workforce
Queen infiltration protocol: kill resident queen, assume her cuticular hydrocarbon profile, achieve colony acceptance through chemical impersonation
Enslaved species: Formica spp., entirely responsible for nest maintenance, foraging, and brood rearing
Assessment: infiltration, assassination, identity theft and proxy labour, a complete colony founding strategy.
Propaganda substances in occupation:
Formica subintegra deploys chemical dispersal agents during raids
Enslaved workers are behaviourally suppressed by the chemistry of their own home, as such workers raised under occupation lack the chemical reference points for resistance
Slave rebellions, what we’ve documented:
Achenbach & Foitzik (2009), Evolution 63:1068–1075: enslaved Temnothorax workers systematically kill 66% of Protomognathus americanus pupae; 83% of future queens destroyed
Method: direct attack (worker gangs dismember developing ants) or selective neglect
Recognition threshold: pupal stage, when species-specific cuticular hydrocarbons become detectable; 95% of larvae survive, lacking discriminable chemical signature
Geographic variation in resistance (Pamminger et al., 2012): West Virginia: 27% pupal survival, New York: 49%, Ohio: 58%
Evolutionary mechanism: kin selection, the rebellion reduces parasite pressure on related host colonies in the surrounding area
Assessment: an organised resistance movement, operating inside the occupied territory, recognised only at the stage when identification becomes chemically possible. Some populations resist harder than others. The variance in resistance is heritable.
1.5 ARMY OPERATIONS: INDUSTRIAL SCALE
Eciton burchellii (New World army ant):
Colony size: 425,000–2,000,000 workers
Raid formation: swarm front up to 200m wide, 200,000 individuals
Operational tempo: statary phase (2–3 weeks, fixed bivouac, pupal development) alternating with nomadic phase (2–3 weeks, new bivouac nightly, maximum advance)
Infrastructure: workers construct living bridges across gaps; fill terrain irregularities with their own bodies; bivouac is built from linked worker chains
Associated species: 350–500+ obligate associates including antbirds (follow columns for flushed prey), parasitic beetles (infiltrate bivouac), hitchhiking mites, specialist spiders
Assessment: the engineering corps is the army. Infrastructure and personnel are the same resource. The camp followers number in the hundreds of species. All the great armies of history generate this economy.
Dorylus spp. (African driver ants):
Colony size: up to 20,000,000 workers
Raid front width: up to 30m
Assessment: the largest army any land organism has ever deployed. On a per-area basis, this exceeds any human military operation in the historical record.
PART TWO: THE CAMPAIGN - LINEPITHEMA HUMILE
The Argentine ant: the century of global conquest
2.1 THEATRE OF ORIGIN
Native range: Río Paraná drainage basin e.g. eastern Bolivia, southern Brazil, Paraguay, Uruguay, northern Argentina
Habitat: riparian floodplain, within a few kilometres of river drainage (Wild 2004)
In-range constraints:
Supercolony size: 25–500m (small)
Intraspecific aggression: high at colonial boundaries
Annual supercolony turnover: ~one-third replaced
Limiting factors: parasitoid phorid flies (Pseudacteon spp.); aggressive competitors; high genetic diversity maintaining effective chemical recognition between colonies
The Argentine ant in its native range is a moderate regional power, constrained by its own internal diversity. Outside of this range the Argentine ant becomes a different beast altogether.
2.2 THE GENETIC WEAPON
The Argentine ant’s global conquest was largely the result of an accident, a founding bottleneck of such severity that it solved the problem of civil war.
Tsutsui et al (2000) established the mechanism: California’s introduced Argentine ant population carried only 23 genetic alleles across 7 microsatellite loci, against 47 in equivalent southeastern US samples. Genetic homogeneity produced uniform cuticular hydrocarbon profiles across nests, these uniform profiles eliminated the chemical basis for colony discrimination. Without the chemical signal for outsiders, a recognition system which had evolved over millions of years to police colonial borders suddenly had nothing to police.
As a result, introduced Argentine ant colonies do not fight each other. Workers from nests a thousand kilometres apart, meeting for the first time, behave as nestmates. The colony no longer has a civil war to fight, thus every unit of aggression is directed outward.
Giraud, Pedersen & Keller (2002) added a further mechanism dubbed “genetic cleansing.” High-density invasive populations actively select against divergent recognition alleles over time, progressively homogenising the chemical landscape. The Argentine ant empire, as it expands, only becomes more unified, not less.
2.3 THEATRE-BY-THEATRE ADVANCE
Maritime cargo of principally coffee and sugar from South America carried Argentine ants to every inhabited continent, at present we can only partially reconstruct this timeline:
Madeira, Portugal (before 1858). The earliest confirmed introduction outside its native range.
New Orleans, USA (1891). Arrived via Brazilian and Argentine cargo vessels.
South Africa (1893). Now split into two distinct supercolonies.
Continental Europe (1890s-1917). Spread from Madeira through Iberia and along the Mediterranean coast.
California, USA (1905-1907). First detected at Ontario, San Bernardino County.
Australia (1939, although possibly as early as 1931). Established successively in Melbourne, Perth, Tasmania, South Australia and Queensland.
New Zealand (1990). Detected at the Commonwealth Games venue, Auckland.
Japan (1993). Entered via Kobe port; spread approximately 900 km along Honshu.
South Korea (2019). Detected at Busan.
Advance rates:
Budding (local spread): ~150m/year where climate permits
Jump dispersal (human transport): 10–360km per event, Suarez et al. (2001)
Conclusion: The Argentine ant campaign of global expansion was opportunistic. Shipping lanes performed the strategic planning, whilst the ants required only the capacity to survive transit in cargo holds and the genetic homogeneity to cooperate with their own kin on arrival.
2.4 CURRENT SITUATION: THE SUPERCOLONY NETWORK
European Main Colony: 6,004km, northwestern Italy to Atlantic coast of Portugal. 30 of 33 sampled populations: no mutual aggression. (Giraud et al 2002)
Catalonian Outlier: Separate supercolony, eastern Spain. Cross-colony trials: lethal combat in 98% of encounters with Main Colony.
California VLC or Very Large Colony: ~1,000km, San Diego to Ukiah. Estimated 30 million combatant deaths annually at southern border.
Australian Network: Melbourne-Adelaide-Perth corridor, ~2,700km. No internal aggression. (Suhr et al, 2011)
Japanese Theatre: Four supercolonies, dominant colony spanning ~900km along Honshu. (Sunamura et al, 2009)
THE INTERCONTINENTAL FINDING:
Van Wilgenburg, Torres & Tsutsui (2010) conducted cross-continental behavioural trials. Workers from the dominant supercolonies of Europe, California, Japan, Australia, and Hawaii were introduced to each other under laboratory conditions. Despite being from the other side of the world, the worker ants groomed each and displayed no aggression. These ants have a shared ancestor who lived in the Río Paraná basin a century earlier, whose lineages had diverged across multiple independent founding events on four continents, yet they recognised each other as nestmates. The authors’ conclusion: “This intercontinental supercolony represents the most populous known animal society.”
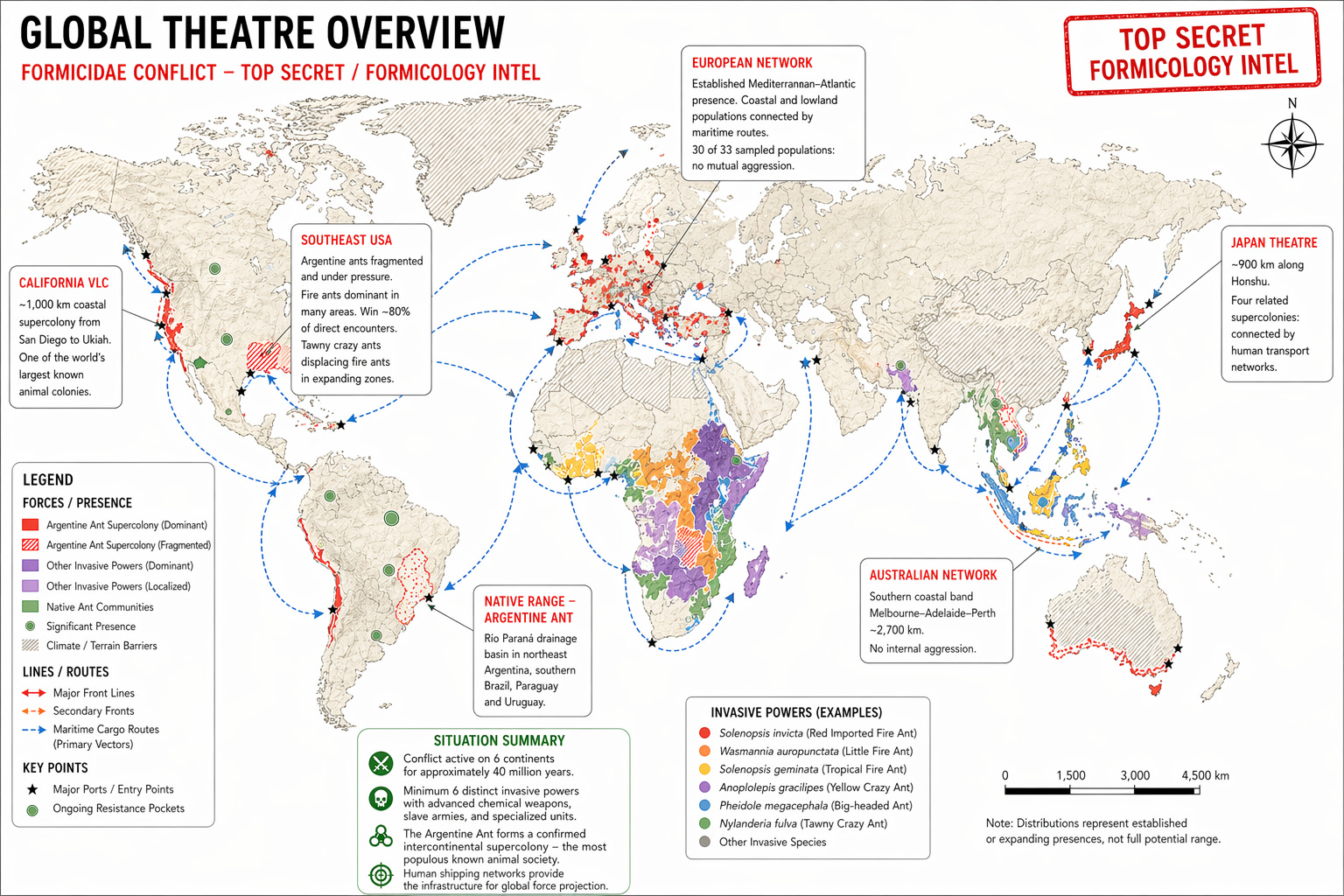
2.5 EFFECTS ON OCCUPIED TERRITORIES
Competitive mechanism upon arrival, effects upon native ants and flora/fauna (Holway, 1999):
Food discovery speed: Argentine ants locate resources in less than half the time of native competitors
Numerical deployment: 4–10× more workers at resources in invaded vs. uninvaded sites
Nest raiding: Argentine ants physically enter and plunder native colonies for brood
California — documented displacement: Pogonomyrmex californicus (California harvester ant), Messor andrei (Harvester ant species), Solenopsis xyloni (California fire ant), Aphaenogaster occidentalis, Formica aerata (Grey field ant), Formica moki, Liometopum occidentale (Velvety tree ant), Monomorium ergatogyna, Tapinoma sessile (odorous house ant). Overall, a near-total collapse of native ant community in riparian habitats (Holway, 1998)
Cascade effects:
Loss of Pogonomyrmex harvester ants > primary prey collapse for coastal horned lizard (Phrynosoma coronatum) > lizard population decline (Suarez, Richmond & Case, 2000)
South Africa: displacement of Anoplolepis custodiens (common pugnacious ant) and Pheidole capensis (seed dispersers) > Proteaceae seeds left unburied > post-fire regeneration failure (Bond & Slingsby, 1984)
Assessment: collateral damage extending three trophic levels beyond the original engagement.
2.6 STRATEGIC LIMITS
The Argentine ant empire has not proven invulnerable, at the edges they have strategic limits created by climatic conditions.
Argentine ants require moisture; cannot penetrate xeric habitats, thus invasion typically limited to ~250m from urban edges in coastal sage scrub (Holway 1998, Bolger 2007)
The 2012-2015 California drought: Argentine ant-occupied area shrank ~30% (fall surveys), ~27% (spring). Native Prenolepis imparis (winter ant) expanded 70% in spring surveys.
Jasper Ridge Biological Preserve long-term monitoring: 26-year decline in Argentine ant occupancy
New Zealand anomaly: Cooling et al. (2012) reported that 40% of surveyed sites showed Argentine ant population collapse. Mean survival time: 14.1 years. Cause: unknown. No human management was involved. To date no competing species identified as responsible.
The empire contracted spontaneously, for reasons researchers cannot perceive, leaving behind communities statistically indistinguishable from pre-invasion baselines. The retreat left no survivors capable of reporting why it happened. Amongst all the documented findings of the Argentine ant projection phase, this is by far the most significant unexplained development. More primary data required.
PART THREE: MULTI-FRONT ANALYSIS
Secondary invasions, overlapping theatres, escalating complexity
3.1 BELLIGERENT REGISTER: SECONDARY POWERS
Red imported fire ant (RIFA) (Solenopsis invicta):
Origin: Pantanal flood plain, northeastern Argentina
US arrival: Mobile, Alabama, 1930’s, via cargo, estimated 9-20 unrelated queens
Global invasion events: minimum 9 independent introductions (Ascunce et al 2011). Mistaken identification may be common, unsure.
European breach: as of 2022 confirmed established population, Sicily (88 nests) (Menchetti et al, 2023)
Combat record: wins 78% of competitive interactions with other ants. 80% against Argentine ants specifically making them a viable global contender
Venom toxicity: 2 to 3× DDT per unit weight, venom contains solenopsin which can cause cardiorespiratory failure in humans
Assessment: RIFA are a power that regularly defeats the Argentine ant in direct confrontation. Currently spreading through all major theatres of war.
Little fire ant, or Electric ant (Wasmannia auropunctata):
Reproductive system: queens produced clonally via parthenogenesis; males clonally via genome elimination; workers sexually as interlineage hybrids
A single founding queen is a complete, self-sufficient invasion kit
Current deployment: Intentional introduction into West Africa as pest control (since 1913). Later invasions into New Caledonia (one supercolony, 450km), Galápagos, Hawaii, Israel, Taiwan, North America, the UK, Cuba and more.
Assessment: little fire ants solved the logistics problem of invasive expansion (requiring a queen plus a founding cohort). A lone queen will suffice.
Tropical fire ant (Solenopsis geminata):
Oldest documented global ant invasion
Vector: Spanish Manila galleon trade, Acapulco to Philippines, 16th century (Gotzek et al., 2015), estimated through genetic data
Assessment: the earliest stages of this conflict predate the modern nation-state. The first recorded supply chain exploitation was the Spanish Empire.
Yellow crazy ant (Anoplolepis gracilipes):
Foraging density: up to 2,254 workers/m², the highest recorded for any ant species (Abbott, 2005)
Assessment: assault capacity and hegemonic power based on overwhelming force of numbers, see Section 1.1 for Square Law combat doctrine
Big-headed ant (US) or Coastal brown ant (AUS) (Pheidole megacephala):
Australian supercolony: no intraspecific aggression across 3000 km (Fournier et al., 2009)
Kenya, 2024: invasion displaced Crematogaster ants protecting whistling-thorn trees, led to increase in elephant browsing and a decline in tree cover. As a result lions switched primary prey from zebra to buffalo (Kamaru et al., 2024)
Assessment: tactical-scale invasions capable of reshaping apex mammal behaviour, ants alter lion prey selection. This exemplar trophic cascade covers approximately four kilometres of savanna food web.
Tawny crazy ant (Nylanderia fulva):
First US detection: Houston, Texas, 2002
Assessment: addressed in full below.
3.2 FRONT LINES: INVASIVE POWERS COLLIDE
The most unpredictable development in the conflict is the emergence of engagements between invasive powers. These forces never encountered each other in their native ranges, none evolved to meet the specific challenges now faced.
The American Southeast: a three-stage displacement
Stage 1: Argentine ants eliminate native ant communities across the southeastern US. Hegemonic regional power for decades.
Stage 2: RIFA established since the 1930’s, then limit and in many areas reverse Argentine ant advance. Fire ants win 80% of direct engagements. Argentine ant populations in the Southeast remain fragmented, with high intraspecific aggression suggesting a smaller, less unified force than their California or European counterparts.
Stage 3: Tawny crazy ants are first detected near Houston in 2002 and identified as Nylanderia fulva in 2012. They displace fire ants across their established range, success coming from their ability to apply formic acid as a solenopsin antidote. This new tactic gives them a huge survival advantage: 98% with the antidote vs. 48% without. The tawny crazy ants achieve densities of 100× all other ant populations combined in peak-density zones. Result: fire ant populations are locally eliminated. LeBrun et al. (2013) reports “most residents prefer the fire ant.” That the liberators proved worse than the occupiers is a situation familiar from the study of human warfare, and has now been observed at insect scale in Texas.
Florida, combined arms: Big-headed ant minor workers engage fire ant workers in holding actions (grasping legs and antennae, preventing effective response) while major workers sever their limbs and follow up by plundering the brood. A two-tier combined-arms operation (1) pin with fast units (2) execute with heavy units.
Bermuda: Argentine ants displace big-headed ants (Wetterer 2017). The same species that lose to fire ants in Florida win in Bermuda. Local conditions (climate, competition history, habitat structure) determine the outcome. There is no absolute universal hierarchy, each species could dominate under the right circumstances.
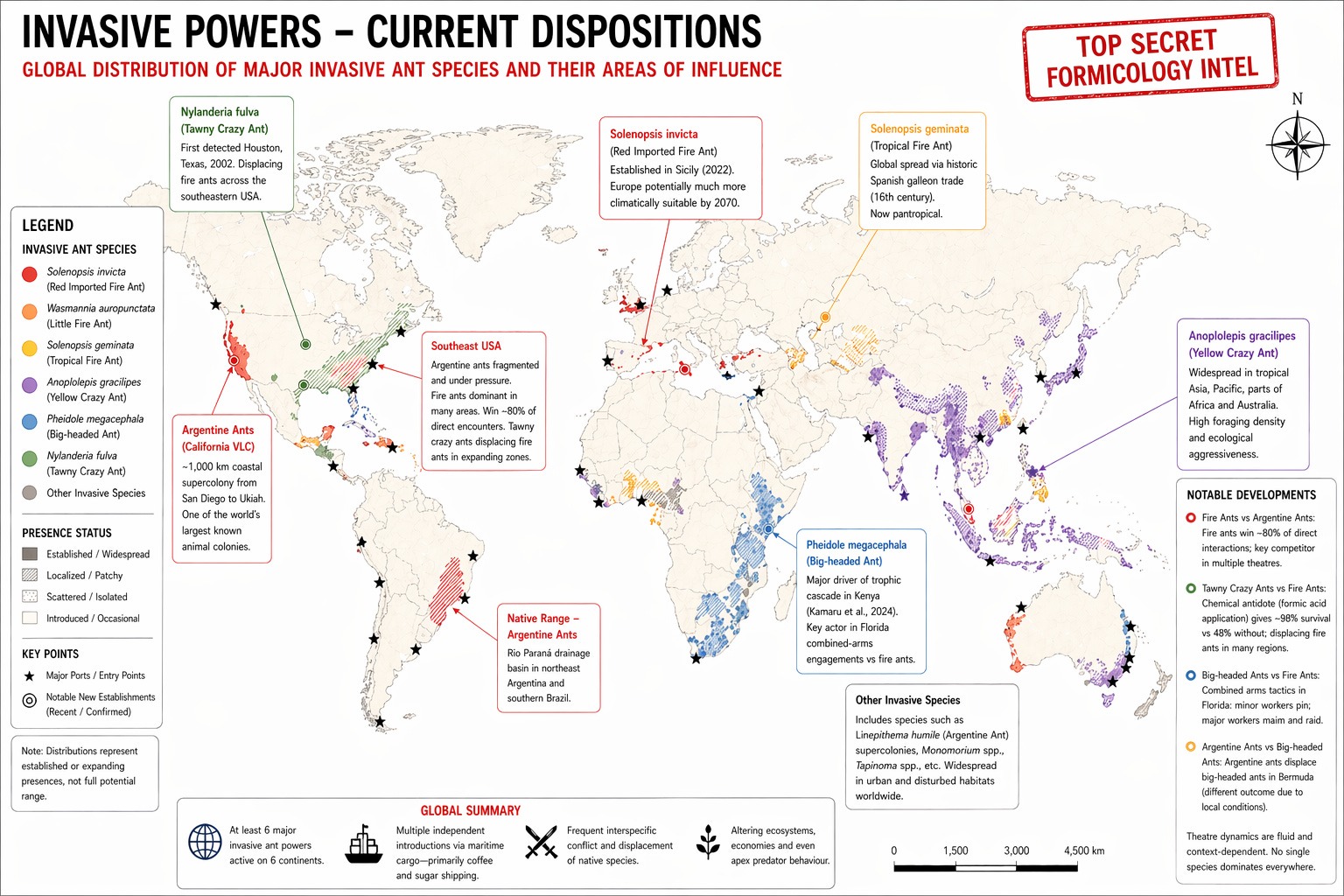
3.3 THEATRE REPORT: CHRISTMAS ISLAND
Christmas Island. 134 km², Indian Ocean. A rainforest ecosystem of unusual isolation, dominated by one keystone species above all others: the red land crab, Gecarcoidea natalis. Pre-invasion population: approximately 43.7–55 million adults. The crabs graze seedlings, turn leaf litter and regulate the structure of the forest floor through sheer biomass. If the crabs are removed, the forest as it exists now would radically change.
Yellow crazy ants (Anoplolepis gracilipes) arrived at some point in the early 20th century and persisted at low density for decades. They were not identified as a threat. The first supercolony was detected in 1989. By 2001, supercolonies covered approximately 25 km², roughly one quarter of all island rainforest, existing at densities of up to 2,254 foraging ants per m². For scale: nest density was 10.5 entrances per m², measuring far beyond a typical infestation.
When the ants encountered the crabs, they sprayed formic acid into the crabs’ eyes and mouthparts. The crabs suffered blindness and dehydration, and died in place, many eaten alive by the ants. Within the supercolony footprint, burrow density collapsed from 1 per m² in intact forest to 0.03 per m², an approximate 42-fold reduction. Millions of crabs were eliminated across a quarter of the island’s surface.
The cascade that followed has virtually no parallel in the ecological record:
Seedling density: 30× higher in invaded zones. With no crabs to graze them, the forest floor underwent impenetrable regeneration.
Scale insects in the canopy: population explosion, tended by ants for their honeydew production. Sooty mould accumulated on the honeydew deposits and led to widespread trees death.
Giant African land snails (Achatina fulica), previously held in check by crab predation: population increase of 253-fold within supercolonies (Green et al., 2011). This secondary invasion was dormant for decades, but was activated by the collapse of the ecological control that had suppressed it.
The academic source for this primary cascade (O’Dowd, Green & Lake 2003) also introduced the concept of an ‘invasional meltdown’, the process by which one invasive species creates conditions that accelerate the establishment and impact of others. Christmas Island is the canonical example in the literature.
The Australian management response was to use aerial fipronil baiting, the campaign began in 2002. Additionally biological control was attempted via the parasitoid wasp Tachardiaephagus somervillei, targeting the ant-scale honeydew mutualism, introduced from 2016. Crab populations show evidence of recovery, but the forest itself has not. Recovery timelines for the canopy die-back zones are unclear.
3.4 RESISTANCE ASSESSMENT OF NATIVE FORCES
Wittman (2014) conducted the most comprehensive review of native ant resistance to invasive species, concluding that the evidence for native ant communities resisting invasive beachheads and overwhelming surges is weak. In suitable habitat, during peak invasion, native species are unlikely to co-occur with foreign armies at meaningful densities.
Documented exceptions:
Prenolepis imparis (winter ant/false honey ant, California) survived the Argentine ant invasion by foraging exclusively in cold months when Argentine ants are inactive. This strategy cannot ultimately defeat the invasion, but it does avoid the worst of it, and prevents total annihilation. Assessment: strategic withdrawal to a niche the invader cannot occupy.
Xeric and high-temperature habitats: Cataglyphis spp. (North Africa), Iridomyrmex spp. (hot inland Australia). Using thermal tolerances which exceed invasive species’ operational range provides another passive defence. Assessment: terrain advantage as last reliable strategic asset.
Island eradication (Channel Islands, California) using human helicopter-dispensed liquid bait, $50,000- 200,000 per km². Feasible only because innate island biosecurity can prevent re-invasion. Assessment: a lasting victory is achievable only where borders can be sealed. On continental landmasses the borders cannot be sealed.
3.5 PROJECTED TRAJECTORY WITH CLIMATE VARIABLE
Bertelsmeier et al. (2015) modelled 15 invasive ant species under 2080 climate scenarios. The counter-intuitive result: the majority were predicted to contract as climatic warming pushes temperatures beyond thermal optima in currently occupied tropical and subtropical zones.
Exceptions with significant implications:
Fire ant suitable habitat in Europe: ~7% of EU territory currently but projected ~30% by 2070 (Menchetti et al 2023). Approximately half of current European urban areas are climatically suitable.
Liu et al (2025) concluded that, across North America, alien and invasive ant species projected to expand range more than native species. This structural divergence favours invaders over the course of this century.
The infrastructure finding:
Bertelsmeier, Ollier, Liebhold & Keller (2017) insisted that, ultimately, human shipping history governs global ant invasion dynamics far more than the climate. Colonial trade routes created invasion corridors that climate models do not capture. A simple takeaway, empires that build shipping lanes also build invasion infrastructure.
CLOSING ASSESSMENT
Returning to San Diego: 30 million annual ant deaths along the borderline represents attrition, a conflict in total stalemate. Neither supercolony is advancing nor retreating, the sheer loss of biomass may eventually grind one or the other into submission, but not yet.
Widening the lens, we have six continents with front lines active for over a century. There are secondary invasions displacing primary invasions in sequences that render the original ecological baseline unrecoverable in human timescales. We have an intercontinental supercolony constituting the most populous cooperative society ever documented on this planet. On one small island in the Indian Ocean we have a complete ecological system dismantled by a species that was simply foraging.
The one force that has demonstrated consistent capacity to reverse the invasion is not a competing species. The climatic forces of heat, aridity, drought: these impartial factors are the only ones with the efficacy to reverse and change the front lines. The New Zealand collapse remains a true mystery, an empire that retreated spontaneously, for unknown reasons, leaving communities indistinguishable from pre-invasion baselines. One generation it was there and fourteen years later, it was not. No successor ant state or invasion has been identified. If there is a mechanism operating in those abandoned sites, it too remains unknown. Working it out could be the most important defensive tool for humans in the coming century, to rout and reverse the flow of invasive ant species.
Beneath the reader’s feet, the engagement continues, as it has done for around forty million years. Your cities, gardens, your patios, terraces, farmlands, fields, your roads, tracks and runways, these may already constitute occupied territory. The warning signs are trail pheromones and alarm compounds and propaganda substances, invasion columns, nest entrances and slave armies. The war began before the human species existed, and will in all probability, continue for long, long after. One day humans may accidentally transport ants to other planets, they may create supply routes that span the solar system and beyond, unintentionally expanding Earth’s most ancient conflict to the stars.
Hells yeah.
It pains me that nobody has taken the task of making an RTS game in this vein. I have a list of like 40 odd ants and their unique traits that would make great upgrade paths or playable races.
There was Ant Rush near 20 years ago, but it was more a coding practice for the dev. Everyone else making ant games are making phone slop.
Hard mode: tyrannomyrmex rex
Now we have an entire article for the Ants are 40k irl meme. Fantastic. I shall read when time allows. Update: that was excellent, the global maps in particular. I would happily read other articles in the same vane regarding the bizarre anomalies nature hides just beneath our noses.